
クラミドモナスやミドリムシは真核生物だが、現代ではもっと原始的な原核生物の古細菌や真正細菌にも光受容の仕組みがある事が分かってきた。高度好塩菌(下写真)は古細菌の一種で砂漠地帯の塩田や塩湖に生息する。

まだ地球上に原核生物しか無い太古の時代には海の塩分濃度が今の5倍も有ったと言われている。https 高度好塩菌はもしかしてその時代に生まれ今より栄えていたのかもしれない。
センサリーロドプシン
高度好塩菌はオレンジ色の光に向かって行く正の走光性と、青緑光や紫光から逃げる負の走光性を持ち、オレンジ光へ向かう傾向と紫光から逃げる傾向はセンサリーロドプシン、青緑光から逃げるのはフォボロドプシンの機能による。その他もっぱら水素イオンを外へ出すバクテリオロドプシン、塩素イオンを細胞内へ取り込むハロロドプシンがある。

走光性を担うセンサリーロドプシンとフォボロドプシンは膜7回貫通型の受容体である点は他のロドプシンと同じだが、膜2回貫通型のトランスデューサータンパク質と結合し複合体を成しているのが特徴である。(下のSRがセンサリーロドプシン、Htrがトランスデューサー)

光受容でレチナールがトランス型からシス型へ変異しロドプシン全体の構造が変化した後、いくつかの中間段階を通ってトランス型に戻る光反応サイクルを形成し、その間にプロトンがタンパク質の中を移動し分子外へ出され、別のプロトンが取り込まれる。
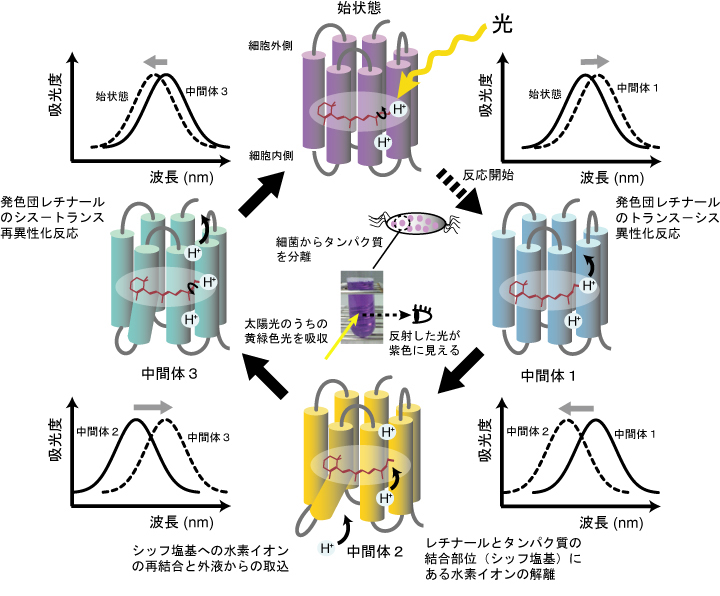
上の図にある様にレチナールは結合しているタンパク質の僅かな変化により吸光波長が遷移するのが興味深い。レチナールだけでは紫外線しか吸収しない。このタンパク質の変化がさらに別のタンパク質へと情報伝達される。高等動物では神経が担う情報伝達がここではタンパク質からタンパク質への情報伝達である。
バクテリオロドプシン
クラミドモナスのチャネルロドプシンがイオンが通過し易い通り道を作り、後は濃度勾配に任せた消極的な「イオンチャネル」であるのに対し高度好塩菌のバクテリオロドプシンはイオンの能動輸送をして濃度勾配を作り出す「イオンポンプ」である。
緑の光に反応して水素イオン(プロトン H+)を細胞外へ出し、細胞の内部を外部の1万倍強い塩基性にする。外へ出た水素イオンは別のタンパク質を通して細胞内へ戻り、その外から内への水素イオンの流れがATPを作るエネルギーとなる。

これは光エネルギーを別のエネルギーに変換する点で光合成に似ているが、光合成の過程よりずっと簡単である。
バクテリオロドプシンやハロロドプシンの様に光エネルギーをATPに変えるイオンポンプと、センサリーロドプシンやフォボロドプシンの様に光エネルギーを情報に変えるタイプがあるが、どうやらイオンポンプの方が原始的でセンサータイプはそこから進化したらしい。https
ここで微生物型ロドプシンについて少し整理してみると、
① タンパク質が色素の吸光波長を制御する。
② 色素のシス–トランスの光異性化でタンパク質の構造変化が起こる事がシグナルとなる。
③ 光を受容したロドプシンはいくつかの中間段階を通って元に戻る光反応サイクルを形成。
④ そのサイクルの中でプロトンの出入りが起こる。
③の光反応サイクルはチャネルロドプシンでも起こっていて(https)恐らく光受容タンパク質全体に共通すると予想される。
④に関してはチャネルロドプシンはプロトンの出入りの代わりにカルシウムイオンやカリウムイオンの出入りがあるので「イオンの出入りによる電位差を作る」点では共通する。

コメント